Высшие растения
Современная наука об органическом мире делит живые организмы на два надцарства: доядерные организмы (Procariota) и ядерные организмы (Eucariota). Надцарство доядерных организмов представлено одним царством – дробянки (Mychota) с двумя подцарствами: бактерии (Bacteriobionta ) и цианотеи, или сине-зеленые водоросли (Cyanobionta).
Надцарство ядерных организмов включает три царства: животные (Animalia), грибы (Mycetalia, Fungi, или Mycota) и растения (Vegetabilia, или Plantae ).
Царство животных делится на два подцарства: простейшие животные
(Protozoa ) и многоклеточные животные (Metazoa).
Царство грибов подразделяется на два подцарства: низшие грибы
(Myxobionta) и высшие грибы (Mycobionta ).
Царство растений включает три подцарства: багрянки (Rhodobionta), настоящие водоросли (Phycobionta) и высшие растения (Embryobionta).
Таким образом, предметом систематики высших растений являются высшие растения, которые входят в состав подцарства высших растений, царства растений, надцарства ядерных организмов.
Отличия высших растений от низших.
Высшие растения отличаются от низших по многим признакам, однако главное их отличие в том, что они по преимуществу - обитатели суши, устроены несравненно более сложно и более разнообразны, чем низшие растения. В отличие от низших растений, высшие растения представляют собой сложно дифференцированные многоклеточные организмы, приспособленные к жизни в наземной среде (за исключением немногочисленных и явно вторичных водных форм), с правильным чередованием двух поколений: полового (гаметофит) и бесполого (спорофит). Переход от гаплоидной ядерной фазы к диплоидной происходит при оплодотворении яйцеклетки сперматовоидом или спермием. И наоборот, переход от диплоидной ядерной фазы к гаплоидной происходит при образовании спор из спорогенной ткани - археспория путем мейоза с редукцией числа хромосом. Таким образом, мейоз у высших растений ведет к образованию гаплоидных спор.
У высших растений имеется сложная система покровных тканей (эпидерма, перидерма, корка), появляется сложный устьичный аппарат. В условиях наземно-воздушной жизни у высших растений появляются мощно развитые механические ткани.
На спорофитах высших растений развиваются многоклеточные спорангии, в которых образуются неподвижные споры, на гаметофитах — многоклеточные половые органы (гаметангии), однако у некоторых голосеменных и у всех цветковых растений гаметангии в процессе эволюции исчезли.
Спорангии одинаковые (равноспоровые растения) или чаще дифференцированы (разноспоровые растения) на 2 типа (микроспорангии и мегаспорангии), гаметангии — всегда двух типов — мужские (антеридии) и женские (архегонии). Как антеридии, так и архегонии защищены многоклеточными стенками. Половые органы высших растений – многоклеточные антеридии (мужские) и архегонии (женские) – взяли начало, вероятно, от многоклеточных гаметангиев водорослей типа диктиоты и эктокорпуса (из бурых водорослей). Женская гамета (яйцеклетка) всегда одна, неподвижна. Зигота у высших растений развивается в многоклеточный зародыш, который проходит первые стадии развития внутри женского гаметофита. Спорофит у высших растений, как правило, расчленён на 3 основных органа — лист, стебель и корень.
В большинстве случаев у высших растений развиваются специальные проводящие ткани — ксилема и флоэма, у некоторых групп редуцировавшаяся. Хлорофилл у высших растений не содержит дополнительных пигментов, окраска фотосинтезирующих частей зелёная.
Происхождение высших растений
Высшие растения, вероятно, произошли от каких-либо водорослей. Об этом свидетельствует то, что в геологической истории растительного мира высшим растениям предшествовали водоросли. В пользу этого предположения свидетельствуют и следующие факты: сходство наиболее древней вымершей группы высших растений - риниофитов – с водорослями, очень сходный характер их ветвления; сходство в чередовании поколений высших растений и многих водорослей; наличие жгутиков и способность к самостоятельному плаванию у мужских половых клеток многих высших растений; сходство в строении и функции хлоропластов.
Предполагают, что высшие растения произошли скорее всего от зеленых водорослей, пресноводных или солоноватоводных. Они имели многоклеточные гаметангии, изоморфное чередование поколений в цикле развития.
Первыми наземными растениями, найденными в ископаемом состоянии, были риниофиты (риния, хорнея, хорнеофитон, спорогонитес, псилофит и др.).
При выходе растений на сушу первыми возникли органы прикрепления
- ризоиды, которые позднее развились в корни. Первые высшие растения обитали в условиях высокой влажности окружающего воздуха, при снижении влажности воздуха возникла необходимость обеспечения растений водой, поэтому ризоиды превратились в корни, поглощающие воду из почвы. Меньшая плотность воздушной среды привела к появлению механических тканей, а необходимость экономить воду – к появлению покровных тканей, а необходимость доставки воды до всех частей растения – к появлению проводящих тканей.
После выхода на сушу высшие растения развивались в двух основных направлениях и образовали две большие эволюционные ветви – гаплоидную и диплоидную.
Гаплоидная ветвь эволюции высших растений представлена отделом моховидные (Bryophyta). В цикле развития мхов преобладает гаметофит, половое поколение (само растение), а спорофит, бесполое поколение, редуцирован и представлен спорогоном в виде коробочки на ножке. Развитие моховидных шло в сторону возрастания самостоятельности гаметофита и его постепенного морфологического расчленения, потери самостоятельности спорофита и его морфологического укрощения. Самостоятельной, вполне автотрофной фазой жизненного цикла моховидных стал гаметофит, а спорофит низведен до степени органа гаметофита.
Мхи как представители гаплоидной ветви эволюции высших растений оказались менее жизнеспособными и адаптированными к условиям жизни на Земле. Их распространение связано с наличием свободной капельно-жидкой воды, необходимой не только для ростовых процессов, но и для полового процесса. Этим объясняется их экологическая приуроченность к местам, где имеется постоянное или периодическое увлажнение.
Вторая эволюционная ветвь высших растений представлена всеми остальными высшими растениями.
Спорофит в наземных условиях оказался более жизнеспособным и адаптированным к разнообразным экологическим условиям. Эта группа растений более успешно завоевывала сушу. Спорофит у них часто имеет большие размеры, сложное внутреннее и внешнее строение. Гаметофит, наоборот, претерпел упрощение, редукцию.
У более простых форм (споровые растения) гаметофит еще имеет самостоятельное существование и представлен автотрофным или симбиотрофным заростком (Lycopodiophyta, Equisetophyta, Polypodiophyta ), а у разноспоровых представителей этих отделов он значительно упрощен, редуцирован. У более организованных – семенных растений – гаметофит утратил самостоятельный способ жизни и развивается на спорофите, а у покрытосеменных (цветковых) сведен до нескольких клеток.
В новых условиях шло постепенное усложнение наземных растений с преобладанием в цикле развития спорофита. Они дали начало ряду самостоятельных групп (отделов) растений, приспособленных к разнообразным условиям жизни на суше.
В настоящее время высшие растения насчитывают свыше 300 000 видов. Они господствуют на Земле, населяют ее от арктических территорий до экватора, от влажных тропиков до сухих пустынь. Они образуют различные типы растительности – леса, луга, болота, заполняют водоемы. Многие из них достигают гигантских размеров (секвойядендрон – 132 м при обхвате 35 м, эвкалипт гигантский – 152 м, вольфия бескорневая – 0,1-0,15 см).
Теломная теория Циммермана
Открытие и изучение риниофитов послужили основой для создания теломной теории, автором которой был Циммерманн. Эта теория раскрывала особенности строения первых наземных растений и показывала возможные пути становления вегетативных и репродуктивных органов высших растений. В основу теломной теории легло понятие телома - цилиндрического участка тела растения, снабженного единственным сосудистым пучком, занимающим терминальное и первично вертикальное положение. Первые сухопутные высшие растения представляли собой, по мнению Циммермана, дихотомически ветвящуюся систему теломов (псилофиты).
В ходе дальнейшей эволюции эта система подвергалась различного рода преобразованиям, которые можно свести к немногим элементарным процессам. К ним относятся "перевершинивание" - смена главной оси, планация - расположение первично трехмерной системы теломов в одной плоскости, изгибание, срастание и редукция.
По мере ветвления теломы из конечных становились промежуточными, поэтому получили название мезомов. Отличительной особенностью теломов было наличие проводящих пучков, построенных по типу протостелы. Этим они отличались от водорослей. Подземные теломы получили название ризомоидов, никаких побегов, а следовательно, и корневищ у первых наземных растений не было. На ризомоидах располагались ризоиды. Вертикальные теломы либо оставались стерильными - вегетативными, либо заканчивались спорангиями. Совокупности теломов нескольких порядков ветвления называются синтеломами; они могли быть вегетативными, спороносными и смешанными. В ходе эволюции теломы превратились в основные органы высших растений - стебли, листья, корни, спорофиллы. Превращение теломов в более сложные органы происходило в результате процессов, протекавших независимо друг от друга. Важнейшие из них - перевершинивание, планация, срастание, редукция. Перевершинивание происходило в результате неравной дихотомии. В итоге формировались главная ось и система боковых ветвей; это явилось предпосылкой для создания крупных форм растений. Под планацией понимается расположение теломов в одной плоскости. Срастание теломов как конечный результат филогенетического процесса мог распространяться на теломы, расположенные как в одной, так и в нескольких плоскостях. Если срастание сочеталось с планацией, то формировались крупные рассеченные листья с системой разветвленных жилок. Срастание теломов, расположенных в разных плоскостях, приводило к образованию стеблей со стелой сложной конфигурации. Под редукцией понимается уменьшение числа порядков ветвления, что приводило к упрощенному строению органа. Так, простая листовая пластинка хвощей в ходе эволюции возникла из системы многократно ветвившихся теломов, которые подвергались редукции. Важным положением теломной теории является положение о том, что возникновение вегетативных листьев и спорофиллов происходило одновременно и независимо друг от друга, и дальнейшее их развитие шло параллельными путями. Следовательно, формирование вегетативных и спороносных побегов происходило также независимо друг от друга.

Рис. 1.Схема эволюции стелы
1 – протостела, 2 – актиностела, 3 - плектостела, 4 – эктофлойная сифоностела, 5 – артростела, 6 – амфифло ная сифоностела, 7 – диктиостела, 8 – эвстела, 9 – атактостела.
В современной флоре протостела характерна для плаунов и некоторых папоротников и встречается в корнях почти всех растений. На поперечном сечении протостела может иметь разную форму. Наиболее примитивным типом является протостела, состоящая из центрального пучка первичной ксилемы, окруженной цилиндром флоэмы. Подобную протостелу называют гаплостелой. Более подвинутым типом является протостела с радиальными выступами ксилемы, называемая актиностелой. Она встречается у некоторых примитивных видов плаунов. Результатом дальнейшей специализации актиностелы является так называемая плектостела, характерная для стеблей большинства видов плауна. В плектостеле ксилема разбита на отдельные пластинки, расположенные параллельно или радиально друг к другу.
В процессе эволюции протостела дала начало сифоностеле. В отличие от протостелы сифоностела имеет трубчатое строение и обладает центральной паренхимной частью, или сердцевиной. Эволюция сифоностелы сопровождается появлением паренхимных участков (лакун) в самом проводящем цилиндре. Благодаря лакунам создается непрерывная связь коры и сердцевины. Различные типы сифоностелы очень характерны для многих папоротников.
Когда сифоностела разбита на сеть или ряды продольных тяжей, получается рассеченная стела, или диктиостела, которая, как и сифоностела, характерна для многих папоротников.
У хвощей и родственных форм из сифоностелы возникает особый тип стелы, названный артростелой. Артростела имеет членистое строение и характеризуется наличием центральной полости и протоксилемных лакун (каринальных каналов). Она характерна для хвощей.
В результате рассечения сифоностелы возникает так называемая эвстела, характерная для большинства семенных растений.
В некоторых случаях, как у ряда травянистых двудольных и большинства однодольных, проводящие пучки эвстелы разбросаны по сердцевине и коре так, что сходство с единой сетью или кольцом теряется совершенно. Ввиду своеобразия подобной стелярной организации она выделяется в особый тип стелы, названный атактостелой. Атактостела развилась в результате крайнего рассечения эвстелы и представляет собой в эволюционном отношении крайне специализированную форму центрального цилиндра.
Происхождение и эволюция листа
Листья возникли впервые у псилофитов и при этом двумя путями.
Микрофиллы (мелкие листья) возникли как выросты (энации) стебля (как у Asteroxylon). Мелкие листья — выросты стебля, постепенно совершенствуясь, дали листья современного характера. Соответственно эта линия развития высших спорофитных растений получила название мелколистной (микрофильной). Сюда относятся плауны, хвощи, некоторые голосеменные, например хвойные.
Крупные листья макрофиллы (мегафиллы), например, листья-вайи папоротников, имели иное происхождение. Присматриваясь к развертывающейся вайе, можно установить, что улиткообразное закручивание пластинки начинается с долек самого последнего порядка, а затем уже последовательно закручиваются дольки высшего порядка, крупные доли и, наконец, вся вайя в целом. Далее оказывается, что каждая долька растет независимо от других своей верхушкой, как стебель. Поэтому, а также учитывая и особенности вхождения сосудистых пучков из стебля в черешок вайи, ботаники пришли к выводу, что лист-вайя папоротника — стеблевого происхождения, уплощенный стебель (ось). Вайя, метаморфизируясь и упрощаясь в пределах самих папоротников, выработала в процессе эволюционного развития листья, которые мы встречаем у высших растений.
Макрофиллы (мегафиллы) возникли в результате срастания теломов типа ринии. Каждый из них имел свою стель. При срастании теломов сливались и их стели. Такие сросшиеся теломы уплощались (кладодифицировались) и превращались в листья-ветви. Такой путь образования макрофиллов можно проследить у одного из псилофитов псевдоспорохнуса (Pseudospornchnus), небольшого (до 3 м высотой) деревца, извест-ного из среднего девона С. Америки.
Папоротниковая линия развития, которую можно вести от псилофитов типа ринии, получила название крупнолистной, макрофильной (макрофиллия — крупнолистность).
До открытия псилофитов оставался большой загадкой вопрос о происхождении высших растений вообще, т. е оставалась зияющая пропасть между слоевцовыми и побегоносными растениями. Вопрос о том, каковы были примитивные сухопутные растения, разрешался лишь в порядке гипотез.
Открытие псилофитов позволило восстановить пути возникновения различных ветвей папоротникообразных и свести к единому источнику разнообразие в строении стелей хвощевидных, плауновидных и папоротниковидных.
Происхождение корня
Тело первых вышедших на сушу растений ещё не было расчленено на побеги и корни. Оно состояло из ответвлений, одни из которых поднимались вертикально, а другие прижимались к почве и поглощали воду и питательные вещества. Несмотря на примитивное строение, эти растения были обеспечены водой и питательными веществами, так как имели небольшие размеры и жили около воды.
В ходе дальнейшей эволюции некоторые ответвления стали углубляться в почву и дали начало корням, приспособленным к более совершенному почвенному питанию. Это сопровождалось глубокой перестройкой их структуры и появлением специализированных тканей. Образование корней было крупным эволюционным достижением, благодаря которому растения смогли осваивать более сухие почвы и образовывать крупные побеги, поднятые вверх к свету. Например, у моховидных настоящих корней нет, их вегетативное тело небольших размеров — до 30 см, обитают мхи во влажных местах. У папоротникообразных появляются настоящие корни, это приводит к увеличению размеров вегетативного тела и к расцвету этой группы в каменноугольный период.
Размножение растений
В процессе онтогенеза растение проходит различные стадии – от молодого состояния к зрелому и в итоге - к гибели. Это закономерный ход развития. Организм непрерывно меняется в процессе роста, в процессе обмена веществ в клетках его накапливаются вещества – отбросы, ненужные организму, изменяющие проницаемость клеточных мембран, химический состав клеток и др. Кроме того, в организме накапливаются изменения, соответствующие изменениям окружающей среды. И как результат этих всех изменений, в зрелом взрослом организме возникают клетки, способные начать жизнь сначала, т.е. организм приступает к размножению. Размножение – одно из обязательных свойств живого организма. Размножением организм разрешает одно из самых сложнейших противоречий – проблему старения и омоложения организма. Механизмы этих процессов пока недостаточно ясны.
Все живые существа имеют давно сложившиеся способы размножения. У растений существуют три способа: 1) вегетативное, 2) бесполое, 3) половое.
При вегетативном размножении дочерняя особь возникает из части тела материнского организма, имеет наследственные признаки и возраст материнского. Вегетативное размножение – это увеличение числа особей данного вида или сорта посредством отделения жизнеспособных частей вегетативного тела растения. При этом размножении обычна и типична регенерация – восстановление целого из части. Способность к вегетативному размножению весьма характерна для растений на всех уровнях организации. Многие нитчатые и пластинчатые водоросли, мицелии грибов, талломы лишайников легко распадаются на части, каждая из которых может стать самостоятельным растением. Это наиболее примитивный способ вегетативного размножения – распад тела без каких-либо специальных органов.
Наиболее разнообразные формы вегетативного размножения наблюдаются у высших растений. У покрытосеменных оно осуществляется частями вегетативных органов (корень, стебель, лист); но особенно часто их метаморфозами: корневищами, клубнями, луковицами, корневыми отпрысками (малина, вишня, яблоня), усами. Эти описанные случаи относятся к категории естественного размножения растений в природе. Они играют колоссальную роль в захвате территории. Все эти способы легко можно использовать для искусственного вегетативного размножения культурных растений. Многие культивируемые растения размножаются также черенками, отводками, путем прививок. С помощью вегетативного размножения человек получает одинаковое потомство и в большом количестве.
При бесполом размножении участвует одна материнская особь. Дочерние особи возникают из одной клетки материнского организма. Т.е. наследуются только признаки материнского организма. Омоложения не происходит или происходит частичное омоложение. Клетки, которые дают начало новому организму, называются спорами. Споры все одинаковые, т.е. не имеют пола. Спора – это клетка для бесполого размножения. Спора – это 1 клетка с ядром, цитоплазмой, запасом питательных веществ (жировые капли), с лейкопластами. Имеет две оболочки: внутренняя – интина, наружная более плотная экзина. Споры по размерам микроскопические, очень легкие, перемещаются ветром, воздухом.
У водных растений (водорослей) споры со жгутиками, подвижные -
называются зооспорами.
Споры формируются в особых органах – в спорангиях (или зооспорангиях – они одноклеточные). Спорангий у наземных растений многоклеточный, имеет стенку, внутри которого находится особая ткань – спорогенная. Клетки спорогенной ткани (имеют диплоидный набор хромосом) делятся мейозом и образуются в огромном количестве гаплоидные споры. Организм, производящий споры и размножающийся с помощью спор, называется спорофитом.
Биологический смысл бесполого размножения: для быстрого и массового расселения и для переживания неблагоприятных условий среды.
У более примитивных высших растений все споры имеют одинаковые размеры и строение. Это явление называется равноспоровость, или изоспория. Из гаплоидной споры вырастает гаплоидное растение, способное к половому размножению – гаметофит. На гаметофите формируются гаметангии (половые органы), и гаметофит – это половое поколение в жизненном цикле растения.

Рис. 1. Жизненный цикл равноспорового растения
Если у растения все споры одинаковые,то гаметофит обоеполый и несет и архегонии (женские гаметангии) и антеридии (мужские гаметангии). Такой гаметофит развивается долго до полного созревания гамет.

Рис.2. Схема жизненного цикла равноспорового растения
У большинства высших растений гаметофит имеет примитивное талломное строение, способен жить только во влажной теплой среде, и при длительном созревании и выживает мало зрелых гаметофитов.
Вероятность оплодотворения сокращается обратно пропорционально времени созревания гамет на гаметофите. Сокращение времени созревания гамет на гаметофите позволяет увеличить вероятность оплодотворения. Направление эволюции высших растений – уменьшение (редукция) фазы гаметофита.
Более организованные высшие растения образуют споры двух видов. Это явление называется разноспоровость, или гетероспория.

Рис.3. Жизненный цикл разноспорового растения

Рис.4 Схема жизненного цикла разноспорового растения
Микроспорой называется мелкая спора высших разноспоровых растений, из которой развивается мужской гаметофит (половое поколение). Мегаспора (макроспора), более крупная из спор, образуемых высшими разноспоровыми растениями, из которой развивается женский гаметофит (половое поколение). Микроспоры, более мелкие, образуются в микроспорангии, а мегаспоры, более крупные, развиваются в мегаспорангиях. Каждая микроспора дает начало одному мужскому гаметофиту, а из каждой мегаспоры образуется по одному женскому гаметофиту. Поэтому некоторые авторы, например Б. М. Козо-Полянский, называют микроспоры «мужскими спорами», а мегаспоры – «женскими спорами». Созревание раздельнополых гаметофитов происходит быстрее, чем обоеполых.
Половое размножение принципиально отличается от вегетативного и бесполого. Половой процесс в растительном мире крайне разнообразен и часто очень сложен, но по существу сводится к слиянию двух половых клеток (гамет) — мужской и женской.
Суть полового размножения или воспроизведения состоит в формировании растением специализированных клеток - гамет (n), в попарном их слиянии (оплодотворении) и образовании зиготы (2n), из которой вырастает новое растение.
Гамета - это половая клетка, ядро которой содержит гаплоидное число хромосом. Сливающиеся гаметы могут различаться между собой по структуре (форме, величине, подвижности), но обязательно физиологически, то есть по полу и по наследственности. В результате слияния гамет в новом организме объединяются материнская и отцовская наследственность. Половой процесс называют оогамия, когда женская гамета крупная, шаровидная, неподвижная (яйцеклетка), а мужская очень маленькая и подвижная (сперматозоид) или неподвижная (спермий). Специализация гамет служит показателем уровня эволюции. Наиболее примитивной формой полового процесса является изогамия, когда обе гаметы одинаковы.
Гаметы формируются в специальных органах, называемых гаметангиями: мужские - в антеридиях, женские - в архегониях. У низших растений гаметангии, как и спорангии, одноклеточные, а у высших - многоклеточные. Архегонии имеют колбовидную форму, антеридии – мешковидную.

Рис. 5. Архегонии и антеридии
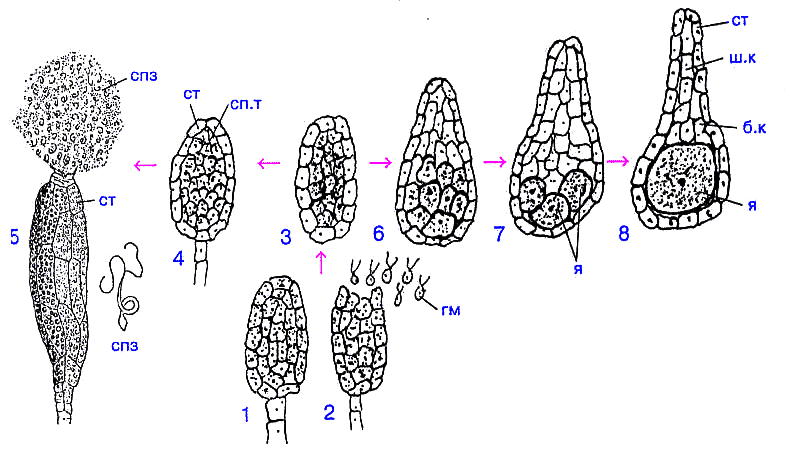
Рис. 6. Происхождение и строение антеридиев и архегониев:
1, 2 - многокамерный гаметангий ;
3 - образование стенки гаметангия;
4, 5 - формирование и строение антеридия;
6, 7, 8 - этапы формирования и строение архегония;
гм - изогаметы; ст - стенка; сп.т - сперматогенная ткань; спз - сперматозоиды ;
ш.к - шейковые канальцевые клетки; б.к - брюшные канальцевые клетки; я - яйцеклетка

Рис.7. Антеридии маршанции.

Рис. 8. Архегонии маршанции
Отдел Моховидные – Bryophyta
Общая характеристика:
Вечнозеленые, автотрофные, в основном многолетние растения;
· Отдел включает 25000 видов;
· Тело состоит из множества клеток и представляет собой либо слоевище (таллом), либо стебелек, от которого отходят боковые побеги, и корнеподобные выросты – ризоидов, корней нет;
· Таллом слабо дифференцирован, но имеются зачатки проводящей и покровной ткани. Механическая ткань в гаметофите отсутствует. В клетках имеются хлоропласты. Они способны к фотосинтезу. Клеточная оболочка состоит из целлюлозы. Глюкозу запасают в виде крахмала. Клетки могут быть диплоидными (2n) и гаплоидными (n).
· Всегда имеется чередование жизненных циклов. Основное поколение – гаметофит (n), спорофит (2n) редуцирован и развивается на теле гаметофита;
· На гаметофите развиваются органы полового размножения: мужские – антеридии, женские – архегонии;
· На гаметофите развиваются органы полового размножения: мужские – антеридии, женские – архегонии. Процесс оплодотворения зависит от воды, так как сперматозоиды подвижны;
· Процесс оплодотворения зависит от воды, так как сперматозоиды подвижны;
· Спорофит является органом бесполого размножения, представлен корневищем, вайями со спорангиями, в которых находятся спорами (n). Споры образуются в результате мейоза;
· После прорастания споры развивается протонема, на которой формируются почки будущих гаметофитов;
· Гаметофиты также способны размножаться вегетативно.
В целом классификация мохообразных выглядит следующим образом:
Отдел Моховидные (Bryophyta)
Класс Печеночники, или Печеночные мхи (Marchantiopsida, или Hepaticopsida)
Подкласс Маршанциевые (Marchantiidae)
Порядок Маршанциевые (Marchantiales)
Порядок Сферокарповые (Sphaerocarpales)
Подкласс Юнгерманниевые (Jungermanniidae)
Порядок Метцгериевые (Metzgeriales)
Порядок Гапломитриевые (Haplomitriales)
Порядок Юнгерманниевые (Jungermanniales)
Класс Антоцеротовые (Anthocerotopsida)
Класс Мхи (Musci, или Bryopsida)
Подкласс Сфагновые, белые, или торфяные мхи (Sphagnidae)
Подкласс Андреевые мхи (Andreaidae)
Подкласс Зеленые мхи (Bryidae).
Класс Печеночники, или Печеночные мхи (Marchantiopsida, или Hepaticopsida)
Подкласс Маршанциевые (Marchantiidae)
Класс объединяет около 6 (10) тыс. видов (300 родов и 50 семейств). Главная особенность Печеночников – дорзовентральное строение тела. В клетках присутствуют особые образования – масляные тельца. Печеночники разделяются на 2 подкласса – слоевищные (Маршанциевые) и листостебельные (Юнгерманиевые), причем последние преобладают по числу видов и распространены в тропиках и субтропиках.
Печёночные мхи — мелкие и нежные мохообразные растения.
Один из наиболее широко распространенных и обычных у нас видов этого класса - маршанция многообразная (Marchantia polymorpha). Это - напочвенное талломное растение в форме дихотомически ветвящейся многослойной дорсивентральной пластинки размером до 10 см, плотно прилегающей к субстрату. Сверху таллом покрыт однослойной эпидермой с разнообразными устьицами, его ассимиляционная ткань разделена на воздушные камеры перегородками. Снизу слоевище обычно несет ризоиды и расположенные рядами эпидермальные чешуйки - амфигастрии.
Талломы раздельнополы, органы полового размножения размещаются на них на особых зонтиковидных подставках, возвышаясь над слоевищем. Мужские гаметофиты имеют волнистые по краю подставки, на их верхней стороне находятся антеридии. Женские прдставки по краю глубоко лопастные. Между долями подставки группами расположены архегонии (шейкой вниз). Они окружены общим покровом - перихецием, а каждый архегонии, кроме того, окружен индивидуальной чашевидной оберткой, называемой псевдопериантием.

Рис.9. Marchantia polymorpha

Рис. 10. Женская подставка маршанции с архегониями

Рис.11. Мужская подставка с антеридиями

Рис. 12. Спорогон маршанции
После оплодотворения из зиготы развивается спорангий в виде коробочки, сидящей на короткой ножке. Коробочка долгое время остается окруженной остатками стенок архегония - калиптрой. Споры в спорангиях при созревании разрыхляются гигроскопичными пружинистыми нитями - элатерами. Прорастающие споры маршанции дают пластинчатую протонему, из которой затем развивается новый гаметофит.

Рис.12а. Маршанция. 1 - таллом с мужскими подставками, 2 - разрез через мужскую подставку, 3 - сперматозоиды, 4 - таллом с женскими подставками, 5 - продольный разрез луча молодой женской подставки, 6 - молодой спорогон, 7 - взрослый спорогон с вскрывшейся коробочкой, 8 - споры и элатеры, 9 - гайстория. сп - споры, к - коробочка, н - ножка, арх - архегоний, ан - антеридий, эл - элатеры.

Рис.13. Жизненный цикл маршанции
Для печеночных мхов характерно вегетативное размножение более или менее специализированными фрагментами гаметофита. Часто оно осуществляется легко отламывающимися верхушками слоевищ или выводковыми почками, образующимися по краю таллома. Когда выводковые почки развиваются в особых органах - выводковых корзиночках, их называют выводковыми тельцами.
Класс Антоцеротовые (Anthocerotopsida)
Класс мохообразных растений, для которых характерны пластинчатые талломы. Представители антоцеротовидных распространены большей частью в тропиках, нескольких видов встречается в регионах с умеренным климатом.
Клетки, из которых состоит таллом, достаточно однородны, для них характерна тонкостенность; масляные тела в них отсутствуют. Верхние клетки содержат хроматофоры с пиреноидами, которые существенным образом отличаются от хлоропластов других высших растений.
В условиях недостаточной влажности у некоторых антоцеротовидных на лопастях таллома формируются клубневидные утолщения, которые сверху покрыты мёртвыми опробковевшими клетками; эти образования могут переносить крайне неблагоприятные условия, а затем прорастать в новые растения.
Большинство антоцеротовидных однодомны; приспособление к недопущению самооплодотворения заключается в том, что антеридии (мужские органы полового размножения) созревают раньше архегониев (женских органов).
После оплодотворения развиваются спорофиты (бесполое поколение); как и у других мохообразных, спорофиты антоцеротовых называются спорогониями. Спорогоний антоцеротовидных состоит из гаустории (всасывающего органа) и длинной узкой коробочки, имеющей форму изогнутого рожка. Стенки коробочки покрыты эпидермой, под которой находится фотосинтезирующая ткань.

Рис. 14. Anthoceros sp.
Класс Листостебельные Мхи (Musci, или Bryopsida)
Подкласс Сфагновые, белые, или торфяные мхи (Sphagnidae)
К сфагновым мхам относятся свыше 300 видов единственного рода сфагнум (Sphagnum) Ветвистые стебли сфагнума усеяны мелкими листьями. На верхушке главной оси боковые веточки образуют розетку почковидной формы. Особенностью сфагновых мхов является непрерывное нарастание стебля верхушкой и отмирание нижней части. Ризоиды отсутствуют, и поглощение воды с минеральными веществами происходит стеблями. Листья этих мхов состоят из двух типов клеток: 1) живых ассимилирующих, длинных и узких, хлорофиллоносных; 2) гиалиновых – мертвых, лишенных протопласта. Гиалиновые клетки легко заполняются водой и долго ее сохраняют. Благодаря такому строению, сфагновые мхи могут накапливать воды в 37 раз больше своей сухой массы. Разрастаясь плотными дернинами, сфагновые мхи способствуют заболачиванию почв. На болотах наслаивание отмерших частей мха приводит к образованию торфяников. Путем сухой перегонки из торфа получают воск, парафин, фенолы, аммиак; путем гидролиза – спирт. Торфяные плиты являются хорошим теплоизоляционным материалом. Сфагновые мхи обладают бактерицидными свойствами.

Рис. 15. Сфагнум
Растение отличается малой теплопроводностью, вследствие чего его часто используют в качестве теплоизоляционного материала в строительстве. Из сфагнума производят дезодорирующие средства, так как он имеет довольно приятный запах. Некоторые народности используют мох данного вида для производства простыней и пелёнок для детей. В цветоводстве сфагнум применяется в качестве наполнителя земляных смесей. Стоит отметить, что в сухом виде мох способен поглощать воду, количество которой в 20 раз превышает массу самого растения. За такие свойства мох пользуется особенной популярностью у фермеров и цветоводов.
Подкласс Андреевые мхи (Andreaidae)
Андреевые мхи – это группа наскальных мхов. Они способны поселяться на монолитной каменистой породе, буквально вгрызаясь в каждую трещинку ризоидами. Формируют довольно плотные дерновинки. В клеточных стенках листьев андреевых мхов находится красный пигмент, благодаря чему растения имеют темно-красный цвет. Этот пигмент выполняет функцию светофильтра для хлоропластов, которые у этих мхов не способны нормально фунционировать при полном освещении.
В мире насчитывается около 100 видов андреевых мхов, большинство из которых произрастают в полярных регионах северного и южного полушарий. Некоторые виды растут в альпийском поясе гор тропиков.Andreidae по некоторым признакам сходны со сфагновыми мхами, по некоторым – с печеночными и бриевыми. Так, их листостебельный гаметофит по внешнему виду напоминает гаметофит кукушкина льна (род Polytrichum). Так же, как и у бриевых мхов, листья имеют центральную жилку. Так же, как сфагновые мхи, андреевые имеют пластинчатую протонему. Коробочка андреевых мхов раскрывается на несколько створок, как у печеночников, но в отличие от последних, верхние концы створок остаются соединенными друг с другом, что придает созревшему спорогону вид японского фонарика. Так же, как и у сфагновых мхов, у андреевых ножка спорогона очень укорочена, так что коробочка выносится вверх с помощью ложноножки, образующейся из гаплоидных тканей архегония.
Подкласс Зеленые мхи (Bryidae).
Насчитывает около 700 родов, объединяющих 14000 видов, широко распространенных повсюду, особенно в тундровой и лесной зонах Северного полушария.
Широко распространен кукушкин лен (Polytrichium commune), образующий густые дерновины на сырых почвах в лесах, на болотах и лугах. Стебли до 40 см высоты, неветвистые, с густыми жесткими и острыми листьями. От нижней части стебля отходят ризоиды. Гаметофиты кукушкина льна раздельнополые. Рано весной на верхушке мужских особей развиваются антеридии, на верхушках женских – архегонии.

Рис. 16. Polýtrichum commúne – кукушкин лен обыкновенный

Рис.16 а. Верхушка мужского гаметофита кукушкина льна с антеридиями

Рис.16 б. Вехушка женского гаметофита кукушкина льна с архегониями

Рис.16 в. Спорогон кукушкина льна

Рис.16 г. Продольный разрез спорогония: 1 – крышечка, 2 – зубцы перистома, 3 – эпифрагма, 4 – стенка урночки, 5 – колонка, 6 – спорангий, 7 – апофиза, 8 – ножка

Рис.17. Кукушкин лен (Polytrichum): 1 - женский гаметофит с архегониями; 2 - коробочка с колпачком; 3 - внешний вид коробочки; 4 - продольный разрез коробочки; 5 - поперечный разрез коробочки; 6 - вскрывшаяся коробочка; 7 - мужской гаметофит; 8 - вершина мужского гаметофита с антеридиями и парафизами; ур - урночка; кол - колонка; спг - спорангий; кр - крышечка; эп - эпифрагма; з.п - зачаточный перистом; п - перистом; кп - колпачок; ан - антеридий; пф - парафизы.

Рис.18. Жизненный цикл кукушкина льна
В природе мхи имеют следующее значение:
1) Они занимают казалось бы непригодные места для жизни;
2) Участвуют в создании биоценоза (особенно в тундре);
3) Именно мох способен накапливать и удерживать радиоактивные вещества;
4) Играют важную роль в природном балансе любых природных зон.
В жизни человека:
1) Мхи могут ухудшать сельское хозяйство, ухудшать продуктивность полей, способствуя их заболачиванию;
2) Предохраняют почву от эрозий (от обсыпания и от отшелушивания ветром);
3) Некоторые виды применяются в медицине;
4) Мхи - источники торфа
Мхи не требовательны к условиям произрастания, они могут поселяться там, где отсутствуют другие растения. Выделяя кислоты, мхи вызывают постепенное разрушение скал, а их отмершие части накапливаются между обломками. Так возникают первичные почвы, на которых со временем поселяются и другие растения. Интересно, что животные почти не питаются мхами. Это способствует накоплению их остатков в почве и, таким образом, увеличению запасов перегноя.
Отдел Плауновидные – Lycopodiophyta
Появление плауновидных связывают с силурийским периодом палеозойской эры. В настоящее время отдел представлен травянистыми растениями со стелющимися, дихотомически ветвящимися стеблями и корнями, а также спирально расположенными чешуйчатыми листьями. Листья возникли как выросты на стебле, и их называют микрофиллы. У плаунов есть флоэма, ксилема, перицикл.
Выделяют два современных класса: равноспоровые Плауновые и разноспоровые Полушниковые.
Класс Плауновые (Lycopodiopsida)
Из всего класса до настоящего времени сохранилось четыре рода.
Род плаун (Lycopodium). К этому роду относят многочисленные (около 200 видов) многолетние вечнозеленые травы, распространенные от арктических областей до тропиков. Так плаун булавовидный (L. clavatum) встречается в травяном покрове хвойных лесов на достаточно влажных, но бедных гумусом почвах. В сырых хвойных лесах широко распространен плаун годичный (L. annotinum). Равноспоровые растения.

Рис.19, Плаун будавовидный

Рис.20. Плаун баранец

Рис.21. Продольный срез спороносного колоска плауна булавовидного (равноспоровое)

Рис.22. Спороносный колосок плауна булавовидного - спорангии со порами

Рис.23. Жизненный цикл плауна булаводиного

Рис.24. Гаметофит плауна

Рис. 24.а. Жизненный цикл плауна булавовидного. 1- спорофит, 2- спорофилл со спорангием, 3 - спора, 4 - гаметофит с антеридиями и архегониями, 5 - молодой спорофит, развивающийся на гаметофите
Класс Полушниковые (Isoetopsida)
Представитель класса род Селагинелла (Selaginella) среди современных родов насчитывает самое большое (около 700) число видов.
Это нежное многолетнее травянистое растение, требующее высокой влажности. Селагинеллы в отличие от плаунов характеризуются разноспоровостью. В спороносных колосках образуется два вида спор – четыре мегаспоры в мегаспорангиях и многочисленные микроспоры в микроспорангиях. Из микроспоры образуется мужской гаметофит, состоящий из одной ризоидальной клетки и антеридия со сперматозоидами. Мегаспора развивается в женский гаметофит, не покидающий ее оболочки и состоящий из мелкоклеточной ткани, в которую погружены архегонии. После оплодотворения из яйцеклетки развивается зародыш, и затем новый спорофит.

Рис.25. Селагинелла безножковая

Рис.26. Спороносные колоски плауна и селагинеллы

Рис. 26 а. Спороносный колосок селагинеллы

Рис. 27. Плаун и селагинелла

Рис.28. Жизненный цикл селагинеллы
Некоторые из сенагинелл живут как эпифиты на деревьях (ведут воздушный образ жизни ― произрастают на стволах или ветвях близлежащих больших растений). Но встречаются среди них и литофиты ― живущие на скальных игорных породах. Однако в большинстве представители этого вида селятся на камнях вокруг рек и водопадов. Размеры растений очень разнообразны, среди них есть виды с 10-ю см по высоте и диаметром стебля только 1 мм (например, селагинелла селавговидная).
Виды плаунов используют как лекарственные, красильные, косметические и декоративные растения.
В научной медицине применяют споры (обычно плауна булавовидного) — прежде в России их называли ликоподий, или плаунное семя — для приготовления детских присыпок, пересыпания пилюль. Споры содержат до 50 % жирного невысыхающего масла, алкалоиды, фенольные кислоты, белки, сахара, минеральные соли.
Заготовку спор производят в конце лета — начале осени, после пожелтения спороносных колосков. Колоски срезают ножницами или острым ножом, обычно в сырую погоду, складывая в мешочки из плотной ткани, затем высушивают на открытом воздухе и просеивают через мелкое сито для отделения спор.
В народной медицине споры плаунов применяют как заживляющее средство для засыпки ран, ожогов, обморожений, при экземах, фурункулах, лишаях, рожистых воспалениях. Стебли используют при заболеваниях мочевого пузыря, печени, дыхательных органов, при недержании мочи, болях в желудке, при геморрое, диспепсиях и ревматизме.
Побеги плауна-баранца применяются как рвотное, слабительное средство, для лечения хронического алкоголизма и табакокурения. Всё растение плауна-баранца содержит ядовитый алкалоид селягин, поэтому лечение должно проводиться под наблюдением врача.
В ветеринарии применяют плауны сплюснутый и баранец, для лечения поноса у коров. Отвар из побегов имеет также и инсектицидное действие, им моют животных (коров, коней, овец, свиней) для защиты от паразитов.
В косметологии плауны применяют при фурункулёзе и против облысения.
Споры также применяют в металлургии для обсыпания форм при фасонном литье — при сгорании их образуется слой газов, препятствующих прилипанию изделия и придающих металлу гладкую поверхность.
В пиротехнике споры иногда добавляют в составы бенгальских огней.

Рис.29. Спороносный колоск селагинеллы (1), 2 - микроспорангий, 3 - мегнаспорангий